从pre-mRNA中去除内含子是由被称为剪接体的大型动态RNA蛋白复合物催化的,该复合物被教科书上称之为细胞里最复杂的超大分子复合物,它是由5个小核糖核蛋白颗粒(snRNPs)和几十个蛋白质因子在每个pre-mRNA底物上从头组装而成的。在剪接反应过程中,多种蛋白-核酸复合物及剪接因子按照高度精确的顺序发生结合、重排和解聚,依次形成预组装复合物U4/U6.U5 Tri-snRNP(U4/U6.U5三小核核糖核蛋白复合物)以及至少8个状态的完全组装剪接体pre-B、B、Bact、B*、C、C*、P以及ILS复合物。其中内含子分支位点(Branch Site,BS)识别是剪接体组装过程中的关键事件。在哺乳动物中,内含子分支位点序列的保守性较差,理论上,仅通过碱基配对机制无法实现精确的内含识别。因此解析出该事件发生时的剪切体形态结构对解决此问题就显得极为重要。
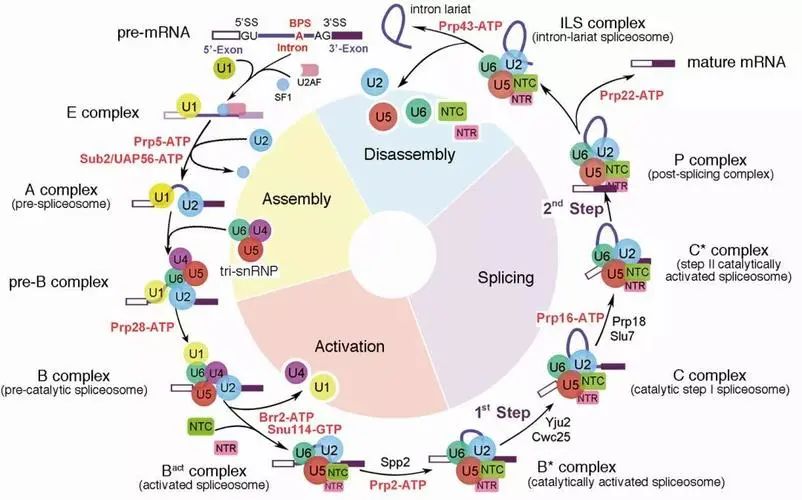
RNA剪接示意图【1】
2021年11月25日,海德尔堡大学的Wojciech P. Galej教授团队在Science上发表了题为Structural basis of branch site recognition by the human spliceosome的文章,在该研究中作者分离了人类17S U2 snRNP,并在体外重构了其依赖于ATP的重塑和与pre-mRNA 底物的结合,从而确定了一系列不同构象状态 U2 snRNP的冷冻电镜结构,包括两个以前未知的组装中间体。这些高分辨率重建提供了对人类U2 snRNP和前剪接体形成的结构和动力学新视角。这些新数据指出HTATSF1和SF3B6在促进pre-mRNA识别方面的关键作用。

为了纯化得到特别代表单一功能状态的U2 snRNP子集,作者使用CRISPR-Cas9介导的基因组编辑将GFP标签引入HEK293F细胞的HTATSF1基因组位点,使用抗GFP的纳米抗体来分离完整的17S U2 snRNP,其中包含U2 snRNA和22种蛋白质,总估计分子量为1.08 MDa。提高分辨率到2.2 Å后,作者发现这些结构与之前的低分辨率结构是一致的。
为了通过冷冻电镜分析获得对BS识别的机制,作者用纯化的17S U2 snRNP和模型BS寡核苷酸(BPS寡核苷酸)在体外重建了BS识别。BPS寡核苷酸与U2 snRNA的27-42位互补,并包括一个凸出的腺苷,用以模拟BP-A。作者通过HTATSF1或DDX46上的GFP标签将17S U2 snRNP固定在抗GFP纳米抗体树脂上,并在各种条件下孵育。在存在ATP和BPS oligo的情况下,U2 snRNP从树脂中释放并在通过甘油梯度离心分析时也能保持与BPS oligo的结合。在没有BPS寡核苷酸的情况下,单独添加ATP也会在升高的温度下诱导HTATSF1解离,这表明这种重构可以在功能上与底物结合分离,从而导致形成重构U2 snRNP。
后续的工作作者解析了这两个新发现的U2 snRNP复合物的高分辨率冷冻电镜结构。
接下来,作者解析了A样U2 snRNP的整体结构。尽管A样U2 snRNP先前的U2 snRNP的低分辨率描述非常一致,但是仍然有跟之前不一致的结构,表明A样U2 snRNP代表了一个不同的剪接中间体。改造后的U2 snRNP与A样U2 snRNP非常相似,除了U2 snRNA 5'端的主要差异以及缺少SF3B6。与17S U2 snRNP类似,这两种复合物都可以分为5'-结构域和3'-结构域,都可以很好地获得结构信息。
在这项工作中,17S U2 snRNP的结构表明HTATSF1和BSL以两种不同的方式相互稳定。BSL和HTATSF1LH/UHM之间可以直接发生相互作用,也可以间接通过U2 snRNA的5'-末端与HTATSF1RRM的可能关联,阻止与BSL竞争的RNA结构的形成。本文的数据表明,模型BPS寡核苷酸可以与U2 snRNP进行碱基配对相互作用。A样U2 snRNP的结构捕获了与分支螺旋相互作用的SF3B6,这提供了两个重要信息。首先,除了SLIIa和SF3A2ZnF之外,它还为U2 snRNA提供了一个特定的结合位点,这使得分支螺旋结合口袋内的U2 snRNA具备了螺旋几何形状。这提供了在缺乏广泛的互补性的情况下一种稳定弱分支点序列的机制。其次,SF3B6在分支螺旋双链体和U2 snRNA的单链区域的连接处结合,因此它定义了分支螺旋的长度和凸起的BP-A相对于其末端的确切位置。经过与BMSL竞争的BS序列将继续通过最近提出的脚趾固定链侵入机制(toe-hold strand invasion mechanism)逐渐形成分支螺旋。该过程中的中间状态(A3'-SSA 复合物)是通过用剪接抑制素A(SSA)阻断剪接体组装而捕获的,在没有SSA抑制的情况下,BS序列将继续完全形成分支螺旋。此时将到达另一个检查点:如果存在凸出的BP-A,它将结合SF3B1中的口袋,这将导致DDX46转变为半封闭构象并解;然而,在没有正确定位的BP-A的情况下,SF3B1HEAT保持开放构象,剪接体停滞。在该复合物中,Prp5为剪接体组装的下一步提供位阻,即募集tri-snRNP。Prp5的长时间阻塞可能会启动丢弃途径。
在这项工作中描述的改造形式的U2 snRNP和pre-A复合物是两种不同的中间体,它们将次优底物引导至丢弃途径。它们代表BS保真度控制中的不同检查点,确保稳定分支螺旋的形成和正确定位的BP-A的存在。
总之,这项工作确定了一系列高分辨率(2.0-2.2 Å)结构,提供了分支位点选择过程的快照。与底物结合的U2 snRNP表明SF3B6稳定了BS:U2 snRNA复合体,这有助于结合序列互补性较差的内含子。
转自网络如有侵权请联系删除 !




